Evolution is most simply defined as descent with modification. Biologists’ understanding of evolution since the 1940s is called the modern synthesis, the synthesis being between evolution through natural selection and the science of genetics.
Population genetics
The modern synthesis employs mathematical (statistical) methods to study what is called a population, the collection of those members of a particular species living in a specified area. Population genetics considers the collective gene pool of such a population, i.e., the sum of all alleles – versions of genes – within the population. A given gene may have more than two alleles, such as those for human blood antigens (A, B and O). The link with what we actually see comes about when these alleles are expressed as phenotypes, i.e., observable traits in the organisms of the species. Evolutionary scientists measure the changes in the distribution (frequency) of the alleles in the gene pool across generations. The underlying genetic composition of the phenotype is the genotype.
The modern synthesis merges genetics and morphology and provides mathematical techniques for a quantitative study of evolutionary change. It recognizes four mechanisms of evolutionary change.
- Natural selection operates when random changes in genes improve the ability of the individuals possessing those genes to survive long enough to reproduce successfully and pass the changes on to their offspring. One then says that differential reproduction has taken place. The superior survival rate of these individuals because of this adaptive trait will assure that, over time, their relative numbers will increase. In terms of population genetics, the frequency of their alleles will increase in the gene pool.
- Mutation is a random change in DNA , the genetic material (much more about that later on). Mutations are small and slow. They may disappear quietly without leading to evolutionary change, or lead to the death of the organism (and the allele). Or they may accumulate very slowly. Mutation is the ultimate generator of variation in alleles and so is necessary for the other three mechanisms.
- Genetic drift occurs when a chance variation (due to mutation) reproduces itself and then gradually becomes important in the gene pool. According to statistics, the range of values of a trait will be well represented by the normal distribution (the famous “bell curve”) in a large population of samples. Since a large population furnishes numerous possible mates, any variation probably will be quickly diluted. Therefore, genetic drift takes place more often in smaller populations which are less statistically significant. Genetic drift occurs most frequently when a sub-group of a population becomes physically separated from the main group, say, due to emigration to an island or environmental isolation by the forming of a new body of water or a mountain ridge – or even just due to large distances between extremities of a vast landmass. The small group may then change sufficiently to form a different species. Such speciation (formation of a new species) due to an external barrier to gene flow is called allopatric, (Greek for “another country”). Since it is due to chance, genetic drift need not be adaptive. Natural selection, on the contrary, is adaptive and so can not bring about evolution of a lasting trait which is bad for the organism.
- Gene flow (or migration) is similar to genetic drift, but involves the immigration of individual organisms or alleles into one population from another. Pollen blown by wind may be an example of gene migration. The result is that genes flow from one population to another.
In all these cases, change is a random, chance occurrence, whereas natural selection is a law which always operates on such changes. In spite of the terms often used to express evolutionary change (“Flowers have colors in order to attract bees.”), it is not teleological (goal-oriented). “Colored flowers attract bees and so survive better.”
Natural selection acts on changes brought about by the other mechanisms cited. In addition, sex is a source of gene mixing, as will be discussed in a later chapter. But the oddest source of change is shuffling of the genetic material when antibodies are created in our bodies. More on that later, too.
Like quantum mechanics, evolution injects the theme of randomness into our understanding of the workings of the universe, but not at all on the same scale.
Species
The notion of species has been much discussed in the history of biology before arriving at the current definition of a species as “groups of interbreeding natural populations, which are reproductively isolated from other such groups”. This means that members of two different groups do not mate to produce viable offspring for one of two reasons: Either (1) they are geographically isolated by necessity or by choice; or (2) their genetic differences are such that they cannot produce viable offspring, or “fertile hybrids”.
However, not much is known about the sexual reproduction of, e.g., bacteria or fossils. So in many cases, biologists must fall back on observation of physical or genetic similarities for distinguishing species. It also happens that observed differences do not amount to differences in species. The case is complicated.
Speciation (formation of new species from an older one) may take place when members of a species occupy different niches or habitats and each group evolves rapidly (in geological terms…) to fit into its respective niche, a phenomenon called adaptive radiation.
In the case of genetic drift, members of the changed smaller group may rejoin the larger one (thus becoming sympatric), bringing their variations with them. The change may take place over a time scale less than that of the dates of adjacent rock strata, in which case there would be no intermediate fossils visible between the two states, hiding any continuity in the evolutionary process. The appearance of evolutionary stasis punctuated by discrete changes has been called punctuated equilibrium, abbreviated as “punk-eek”.
Evolution can give rise to two kinds of similar structures. Characters of different species which are inherited from a common ancestor are called homologous. An example is the presence of the four limbs of tetrapods. Similar characters which are not inherited from a common ancestor are analogous, like the wings of bats and birds, and are the result of convergent evolution, the evolution of similar adaptations in species which are at most distantly related.
Fossils and classification
The goal of evolutionary classification is to demonstrate and explain relations among species. The method uses hierarchical grouping of species into larger groups called genera (singular, genus), genera into families into orders into classes into phyla into kingdoms. This amounts to a road map or genealogy of evolutionary relations. Since two species may be related by being descendants of a long-extinct ancestor, information about that extinct ancestor is necessary. This is obtained primarily through the evidence of fossils.
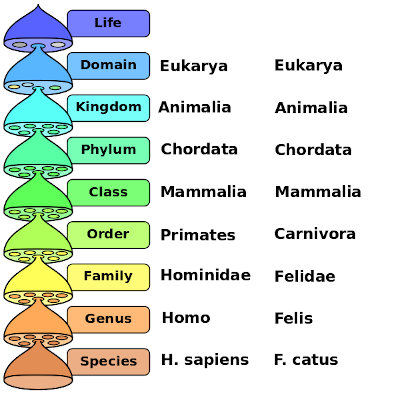
Classification of modern humans and house cats, after Wikipedia
These classifications will be discussed in more detail in following chapters.
Fossils and fossilization
Fossils are central to our understanding of past species, but they come with problems.
How fossils are formed
If a dead organism does not decay and is not destroyed by predators, it may be covered, all or partially, by sediments. The sediment continues accumulating and, under pressure, eventually may form rock. Shells may dissolve and leave a hole in their own form in the sediment – a mold. Dissolved minerals such as silica may move through the porous rock, fill in the hole and harden, taking on the shape of the organism. Or the dissolved minerals may fill in only the pores in the organism itself, leaving a detailed record. The result is a fossil, entombed within the sedimentary rock.
Fossils may also be found where organisms are frozen (as in Siberia) or dessicated (as in deserts). They may be preserved in amber from pine resin or in tar if they fall into a petroleum swamp. They may leave only impressions, tracks or even footprints which are then preserved, for instance, by falling ash.
From the number of species alive today and the “turnover” seen in fossils, an estimation of the number of species which have existed during Earth’s history gives numbers in the billions. But the fossil record only contains some hundreds of thousands, so the fraction of the number of past species in the fossil record is much, much less than 1%. Where have all the fossils gone?
What can fossilize
Some fossils have indeed gone, but many others were never formed. Organisms consist of parts of differing hardness. Teeth are the hardest and decay slowly enough that they may be fossilized easily. Bones decay faster than teeth, but still slowly enough to be fossilized. However, soft body parts usually decay before adequate sedimentation can take place. Only if sedimentation occurs rapidly can soft parts be preserved, as in the case of the mudslide which preserved the fossils of the Burgess Shale and those of Chengjiang, CHiina..
Many fossils are unable to form because of their environment. The ideal habitat for fossilization is a shallow basin which is slowly subsiding, providing good conditions for marine animals and for an appropriate sedimentation rate. This is no help for terrestrial animals.
Even if fossils are formed, they may be destroyed by subsequent movements of the earth, during which heat and pressure may deform, break, crush or burn them up. No fossils are found in igneous rocks and almost none in metamorphic rocks.
Fossils are often hard to find. Most of the earth has not yet been searched for fossils. They often are found when their enclosing sediments erode or are exposed by geological activity, such as at the East African Rift. Such conditions do not take place everywhere.
For all these reasons and more, the fossil record is a very incomplete history of life on Earth.
Classification – taxonomy and cladistics
According to the theory of evolution, all living organisms of one species have evolved from organisms of a different species, and those from still different species, all the way back to the original Ur-form of life, the first cell or amino acid or whatever it was.
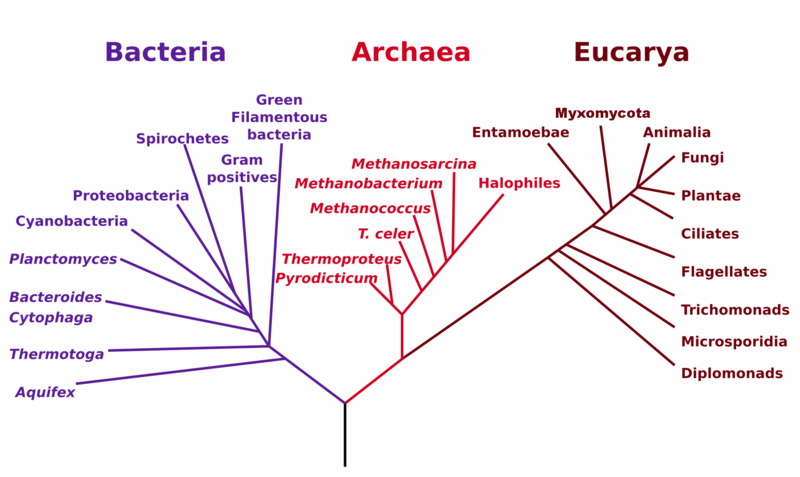
The branch of biology called taxonomy is the science of defining groups of organisms based on shared characteristics in order to show up their shared evolutionary history. The result may be displayed as a family tree or phylogeny. Biologists use cladistics, or phylogenetic systematics, to diagram the evolutionary steps between species. An example of such a diagram is in the above figure. A clear distinction is made between primitive characters and advanced or derived characters. Only groups defined by common derived characters are considered valid monophyletic groups or clades, which are groups descended from a common ancestor and which include all descendants of the common ancestor. Monkeys and apes without humans do not constitute a monophyletic group, nor do reptiles without birds. It is important to understand that a phylogeny is a tree, not a ladder, and implies nothing about whether an organism is “advanced” or “primitive”.
Although these phylogenies are based on both morphological and molecular data, new data often leads to slightly different relations, so the tree changes somewhat as time goes by. This is due mainly to the incomplete record presented by the fossils.
Next, atomic physics and chemistry.


{kind=link}